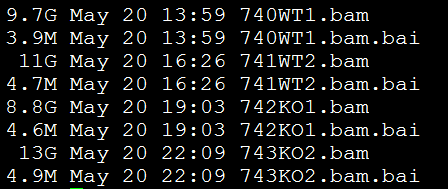
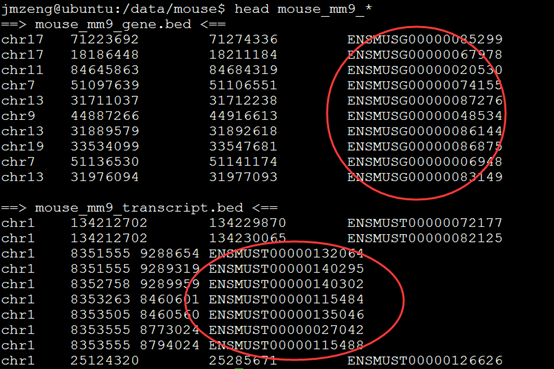
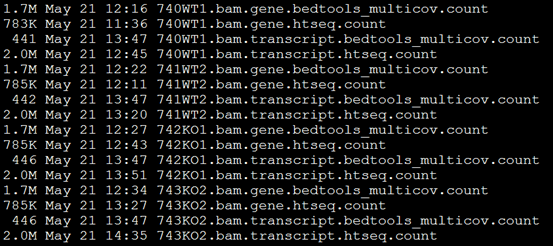
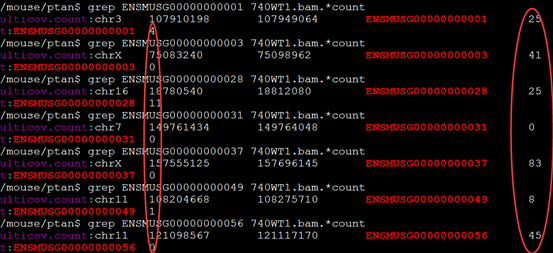
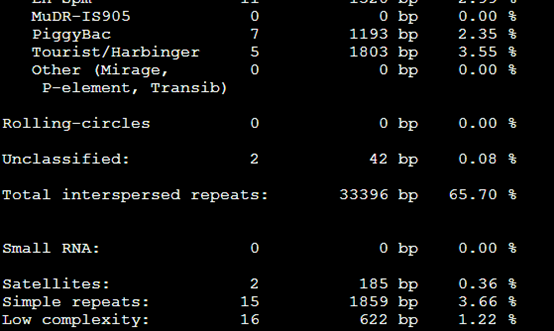
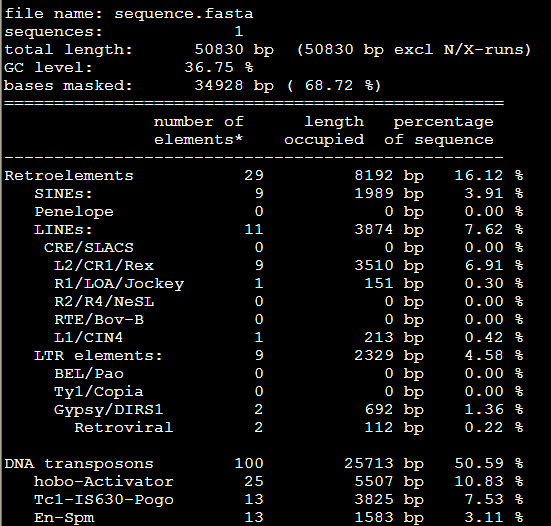
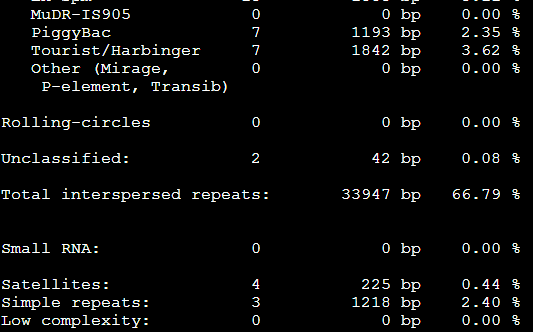
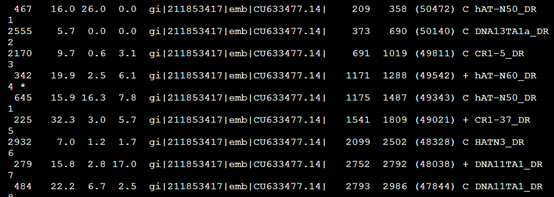
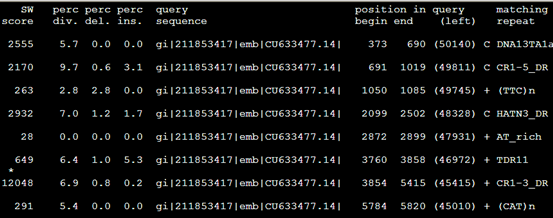
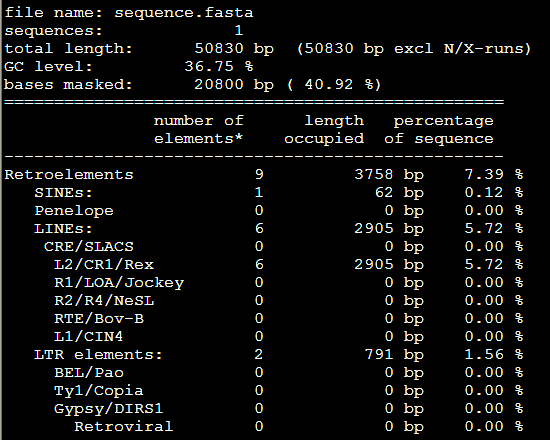
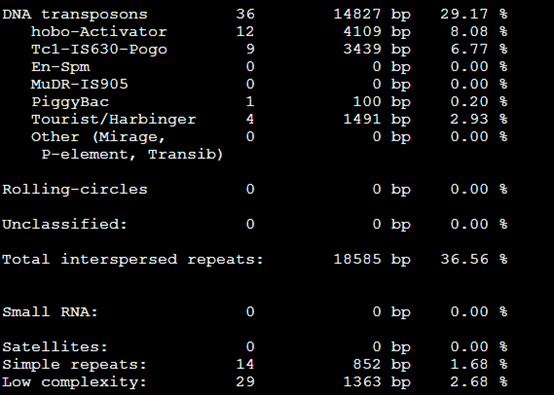
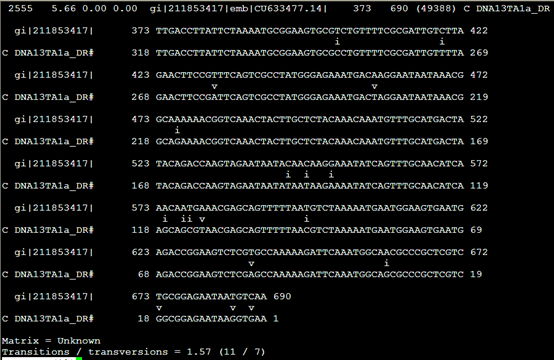
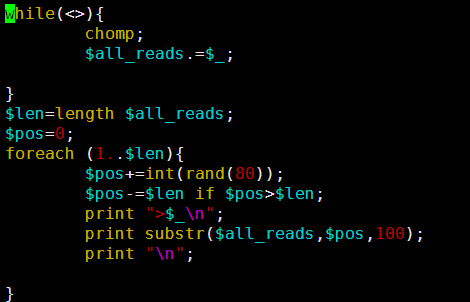
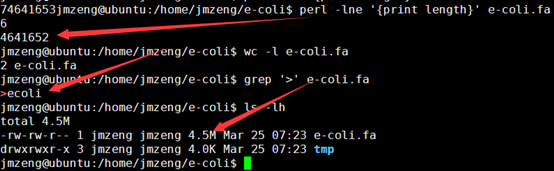
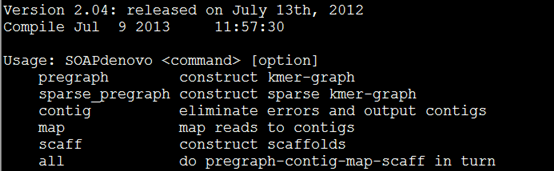
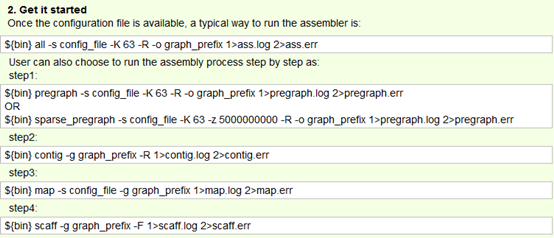
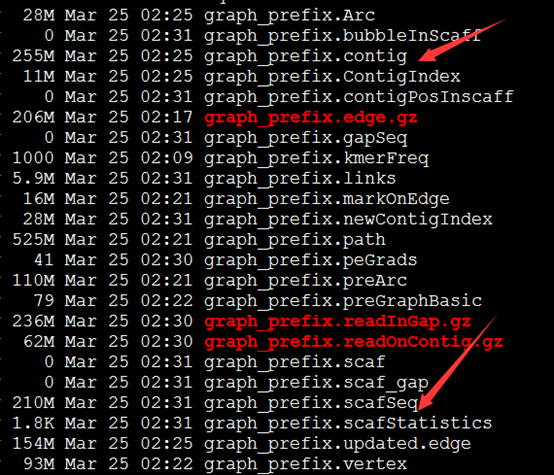
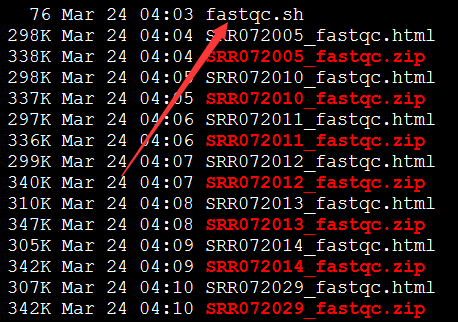
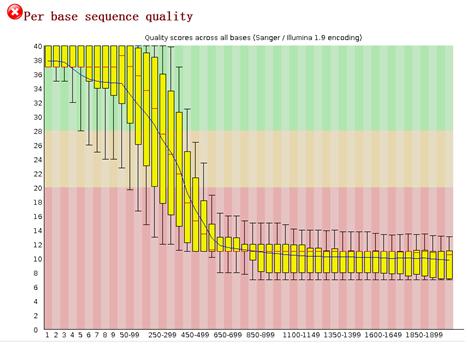
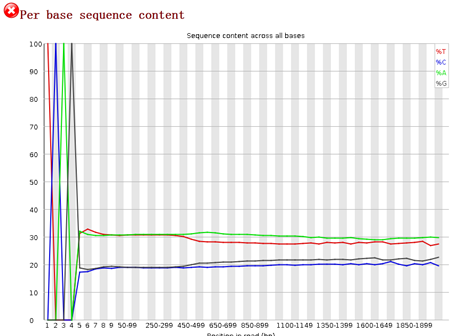
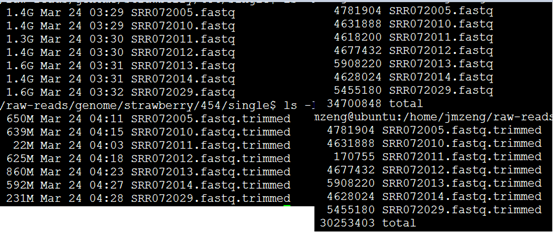
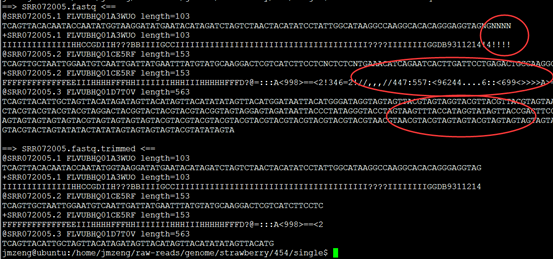
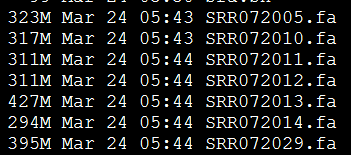
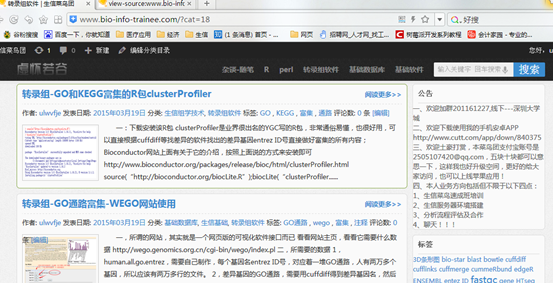
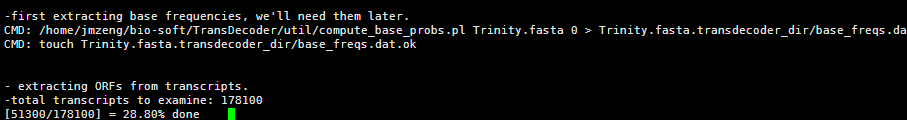
要保证当前文件夹下面有了742KO1.count等4个文件,就是用htseq对比对的bam文件进行处理后的输出文件
library(DESeq)
#加载数据
K1=read.table("742KO1.count",row.names=1)
K2=read.table("743KO2.count",row.names=1)
W1=read.table("740WT1.count",row.names=1)
W2=read.table("741WT2.count",row.names=1)
#列名
data=cbind(K1,K2,W1,W2)
#如果是htseq的结果,则删除data最后四行
n=nrow(data)
data=data
[c language="(-n+4:-n),"][/c]
#如果是bedtools的结果,取出统计个数列和行名
kk1=cbind(K1$V5)
rownames(kk1)=rownames(K1)
K1=kk1
#差异分析
colnames(data)=c("K1","K2","W1","W2")
type=rep(c("K","W"),c(2,2))
de=newCountDataSet(data,type)
de=estimateSizeFactors(de)
de=estimateDispersions(de)
res=nbinomTest(de,"K","W")
#res就是我们的表达量检验结果
到这里,理论上差异基因的分析已经结束啦!后面只是关于R的bioconductor包的一些简单结合使用而已
library(org.Mm.eg.db)
tmp=select(org.Mm.eg.db, keys=res$id, columns=c("ENTREZID","SYMBOL"), keytype="ENSEMBL")
#合并res和tmp
res=merge(tmp,res,by.x="ENSEMBL",by.y="id",all=TRUE)
#go
tmp=select(org.Mm.eg.db, keys=res$ENSEMBL, columns="GO", keytype="ENSEMBL")
ensembl_go=unlist(tapply(tmp[,2],as.factor(tmp[,1]),function(x) paste(x,collapse ="|"),simplify =F))
#为res加入go注释,
res$go=ensembl_go[res$ENSEMBL]#为res加入一列go
#写入all——data
all_res=res
write.csv(res,file="all_data.csv",row.names =F)
uniq=na.omit(res)#删除无效基因
sort_uniq=uniq[order(uniq$padj),]#按照矫正p值排序
#写入排序后的all_data
write.csv(res,file="all_data.csv",row.names =F)
#标记上下调基因
sort_uniq$up_down=ifelse(sort_uniq$baseMeanA>sort_uniq$baseMeanB,"up","down")
#交换上下调基因列位置
final_res=sort_uniq[,c(12,1:11)]
#写出最后数据
write.csv(final_res,file="final_annotation_gene_bedtools_sort_uniq.csv",row.names =F)
#然后挑选出padj值小于0.05的数据来做富集
tmp=select(org.Mm.eg.db, keys=sort_uniq[sort_uniq$padj<0.05,1], columns="ENTREZID", keytype="ENSEMBL")
diff_ENTREZID=tmp$ENTREZID
require(DOSE)
require(clusterProfiler)
diff_ENTREZID=na.omit(diff_ENTREZID)
ego <- enrichGO(gene=diff_ENTREZID,organism="mouse",ont="CC",pvalueCutoff=0.05,readable=TRUE)
ekk <- enrichKEGG(gene=diff_ENTREZID,organism="mouse",pvalueCutoff=0.05,readable=TRUE)
write.csv(summary(ekk),"KEGG-enrich.csv",row.names =F)
write.csv(summary(ego),"GO-enrich.csv",row.names =F)